Starch
.........................................................................................
Introduction
Starch is produced by green plants for energy storage over long periods. It is synthesised in a granular form in special organelles, plastids, as two principles types. In chloroplasts a temporary storage form is produced during photosynthesis, whereas long-time storage starch is produced in amyloplasts. Starch granules are mainly found in seeds, roots and tubers, as well as in stems, leaves, fruits and even pollen. Grain seeds, such as maize kernels, contain up to 75% of starch. Biosynthesis of starch granules is initiated at the hilum, and the granule grows by apposition. The granules occur in all shapes and sizes (spheres, ellipsoids, polygon, platelets, and irregular tubules). They have diameters ranging from around 0.1 to 200 mm depending on their botanical origin  . Generally the difference in external morphology is sufficient to allow unambiguous characterization of the botanical origin, throughout optical microscopy.
. Generally the difference in external morphology is sufficient to allow unambiguous characterization of the botanical origin, throughout optical microscopy.

Fig. 1 Representations of some typical morphologies of starch granules
Starch granules are densely packed with semi-crystalline structures, the crystallinity varying from 15 to 45%, the density being about 1.5 g/cm3. In the granular form, starch can be easily isolated by gravity, sedimentation, centrifugation and filtration. It can be subjected to various chemical, physical and enzymatic modifications with subsequent washing and processing.
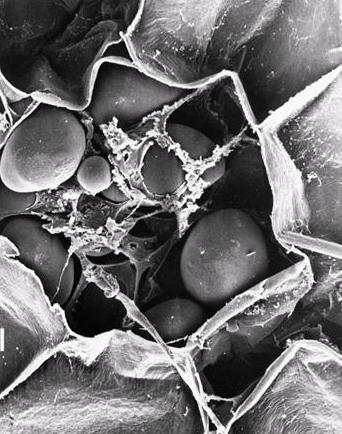
Most of the native starch granules exhibit a Maltese cross when observed under polarized light. The positive birefringence indicates a radial orientation of the principle axis of the crystallites. However, birefringence remains unchanged on both polar and equatorial sections of elongated starch granules, indicating that crystallites are extremely small and exhibit multiple orientations, which interferes during observations. Native granules yield X-ray diffraction patterns of low quality, as the bulk of the starch polymers are in an amorphous state (70 % on average).
 ____
____ 
Fig. 2 Representations of a birefringent starch granule
Starch is a biopolymer and consists of two major components: amylose and amylopectin. Amylose that builds up to 15-35% of the granules in most plants, is a primarily linear polysaccharide with α-(1→4)-linked D-glucose units. Some amylose molecules, particularly those of large molecular weight may have up to 10 or more branches.
Amylopectin, is a highly branched molecule, with α-(1→4)-linked D-glucose backbones and exhibits about 5% of α-(1→6)-linked branches, which have a profound effect on the physical and biological properties.
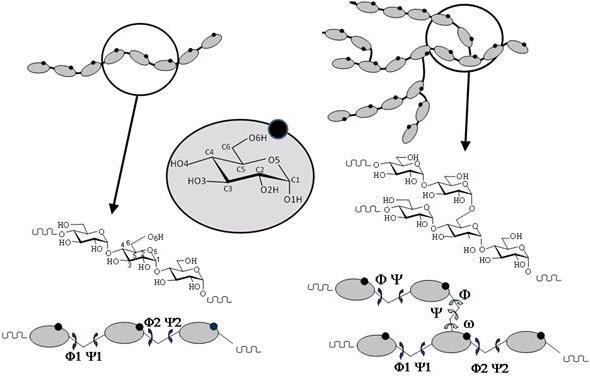
Fig. 3 Schematic representation of amylose and amylopectin
At the lowest level of structure, most starch granules are made up of alternating amorphous and semi-crystalline shells which are between 100 and 400 nm thick . . . . These structures are termed “growth rings”. The radial organization of the amylopectin within such structures is thought to cause optical polarization, since the visible optical polarization is in the order of the wavelength of the visible light (100 to 1000 nm). At a higher level of molecular order, X-ray diffraction investigations indicate a periodicity of about 9 to 10 nm within the granule. . . . The periodicity is interpreted as being due to crystalline and amorphous lamellae found within the semi-crystalline shells. These would be formed by clusters of side chains, branching off from the radially arranged amylopectin molecules. These would appear to be a universal feature in the structure of starch, independent of botanical source. Furthermore, it suggests a common mechanism for starch deposition . Good quality powder diffraction patterns can be obtained from starch powder treated by mild acid hydrolysis to remove amorphous materials. These patterns can be used to identify several allomorphs and to group most starches conveniently according to their physical properties. Generally, most cereal starches give the so-called A-type pattern, some tuber starches (such as potato) and cereal starches rich in amylose yield the B-pattern; legume starches generally have a C-type pattern.
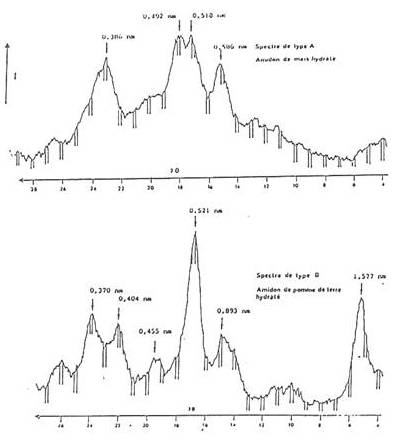
Fig. 4 Typical X-ray powder diffractograms of A abd B starch
Single Helical Structure of Amylose
In a freshly prepared aqueous solution, the amylose chain adopts a random coil structure, which is, however, not stable . Amylose tends to form, in an instant reaction, single helical inclusion complexes with suitable complexing agents . The single helical (inclusion) complex was first reported by Katz and van Itallie . After cooking, dough develops an X-ray diffraction pattern which differs from the original A, B or C patterns that was named V-pattern from the German word “Vaklinestorone” meaning gelatinization .

Fig. 5 Representation of the 3D structure of amylose-V
repeat unit
Double Helical Structure of Amylose
In the absence of complexing agents, amylose molecules gradually associate with a rate that depends on the molecular size
. A minimum chain length of DP 10 is required for double helix formation in a pure oligosaccharide solution
. Shorter oligosaccharides such as maltohexaose can co-crystallize in the presence of longer chains . These results indicate that DP 6 is the shortest branch chain-length found in amylopectins of all native starches.
Structures of retrograded double helical amylose have been investigated by X-ray fiber diffraction methods using methodology summarized in
.
They display A- and B-type X-ray diffraction pattern . Similar observations we made for the single crystals grown in vitro from mono-disperse fractions of amylose having DP 15 and DP 30, which give rise to a powder diffraction pattern typical of the A-type and B-type, respectively.

Fig. 6 Representation of a X-ray fiber and electron diffraction pattern of A and B starch
The three-dimensional structures of the crystalline arrangements corresponding to the A-type and the B-type have been elucidated throughout the joint use of X-ray fiber diffraction, electron diffraction and molecular modelling and crystallographic study of small oligosaccharides . The crystal structure of A-type starch has been revisited using synchrotron radiation microdiffraction data collected from individual micron-sized single crystals analysis of single crystal. Whereas all the main features of the original structure determination are corroborated, this new investigation provides the unambiguous location of a new water molecule bringing as 1.622 the calculated density of the A-type crystal .
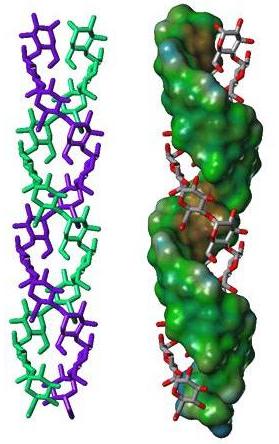
Fig. 7 Representation of a 3D structure of A starch double helix and the projection of the structure
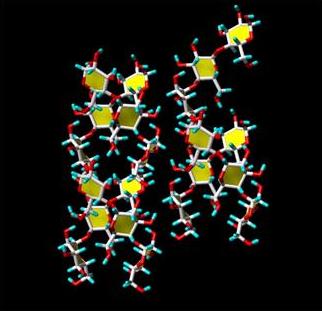
The double helices in both A- and B-type are left-handed and parallel stranded. They are almost perfectly six-fold structure, with a crystallographically repeating unit of 10.5 Å. The symmetry of the double helices differs slightly in A- and B-structure. In the A-form, the repeated unit is a maltotriosyl unit, whereas a maltosyl unit makes up for the repeat of the B-type. Independent evidence about the nature of these repeated units have been gained from solid state 13C-NMR, as the C-1 peak in the A-form is a triplet, whereas it is a doublet in the spectrum of the B-form . In the A- and B-allomorphs, the observed space group imposes the parallel arrangement of all double-helices. The double helices of both forms are packed in hexagonal or pseudohexagonal arrays. The void in the lattice of B-starch, which accommodates numerous water molecules, is not present in A-starch. In both arrangements, there is a pairing of double helices that corresponds to 1.1 times the distance between the axes of the two double helices. A relative translational shift of 0.5 nm along the orientation of the chains allows a very close nesting of the “crests” and “troughs” of the paired double helices. Such a dense association, which is strengthened by O-2...O-6 and O-4...O-3 hydrogen bonding, corresponds to the most energetically favored interactions between two double helices, as shown by theoretical calculations . Therefore, the observed crystal structure of both polymorph A and B of crystalline starch can be rationalized, based on energy calculations, and key structural components have been identified. Among all possible arrangements, only two appear to be sterically favoured. These two types of arrangements of double helices are found in the A-polymorph. The B-polymorph is based on only one type that is favoured by the duplex of parallel double helices. These calculations could rule out the possibility of forming another stable three-dimensional arrangement, and consequently the occurrence of the C-polymorph.
 ___
___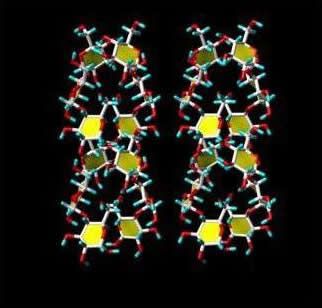
Fig. 8 Representation of a 3D structure of the two types of duplexes of starch double helix
The structural characteristics of the branching point of amylopectin are difficult to assess, however, since they are thought to be located in the amorphous regions between crystallites and since they constitute such a small fraction of the total material. Some basic structural features have, however, been established through computer modeling
. In particular, it was found that among the low energy arrangements, the side chains would fold back onto the carrying chain, thereby producing dense three-dimensional structures in which a parallel arrangement is achieved.
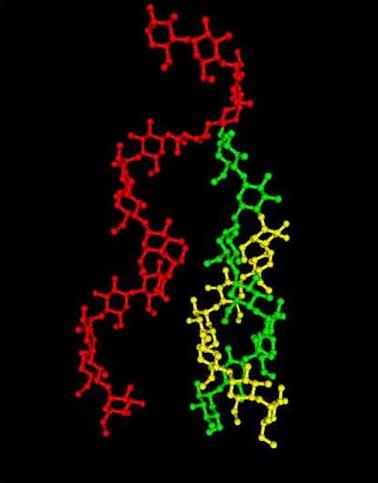
Fig. 9 Representation of a 3D structure of α-1→6 linked chain branched on a single chain (and/or double chain)
Furthermore, the branching between two strands of the double helices was investigated. It was found that one particular set of conformations about the glycosidic linkages in the two different strands could result in an arrangement such that double-helical strands could be connected through a α-(1→6)-linkage, with a minimum of distortion . This indicates that the branch points do not induce extensive defects in the double helical structure, but instead may serve to initiate the crystalline arrangement.
The concept of amylopectin forming double helices easily integrates into the currently accepted cluster model, with the short linear chains of the branches being intertwined into double helices, whilst the α-(1→6)-branch points are located in the more amorphous regions between the clusters of double helices. This concept helps in understanding that parts of the amylopectin molecules are capable of forming double helices, which explains the apparent anomaly that a branched polymer is the source of structural order within the granule.
According to Hizukuri , amylopectin molecules in A-type pattern starches have shorter constitutive chains and more of the short-chain fraction than amylopectin molecules in the B-type starches. Jane et al., determined that A-type starches had branch points scattered in both amorphous and crystalline regions, although for its own part, the B-type starch had most branch points clustering in the amorphous ones. It was concluded that the branching pattern of amylopectin played a key role in the determination of the type of crystallinity. Such point of view was confirmed throughout an investigation of the role of the distance between branching points in a cluster related to crystallization pattern of the chain arrangement . The study was performed on two allomorphs, double maize mutants of A- and B-type (wxdu and aewx, respectively). It was shown that the branching zone of clusters in A-type clusters was larger but with shorter distance between the branching points than in the B-type branching zone of clusters. These results indicate that both chain length specificity combined with α-(1→6)-linkage distribution, have to be considered in the cluster feature.
A further interesting observation regarding starch granule molecular order has been gained by solid state 13C NMR investigation of starch . The ratio of single to double chains in a given starch sample can be assessed. The investigation suggested that the level of helical order is often significantly greater than the extent of crystalline order; which means that starch granules contain double helices moieties, which are not involved in extended crystalline arrays. A novel approach for calculating starch crystallinity and its correlation with double helix content has been developed based on the joint use of X-ray diffraction and solid NMR . The results indicate that a significant proportion of double helices present in starch granules are pack into somehow imperfect crystalline structures. It would therefore appear that this observation is relevant to the amylopectin component of the granule (since amylose is not thought to be extensively in the double helical state); it suggests that much of the amylopectin in the more amorphous shells of the granule is in the double helical form. As mentioned above, computer modelling supports such a view that double helices in amylopectin are sustainable in terms of molecular energetic, even at the α-(1→6)-branch points located in the amorphous regions.
Advances in the field of molecular modelling offer some new possibilities to investigate further the nature and importance of both branch points and amorphous/crystalline interfaces in starch. In approaching these questions, it is wise to adopt the attitude that one is not searching for single answers, but for all of the indications that would lead to a clearer image of how starch macromolecules are arranged and interact in nature. To this end, subunits of single helices, double helices and branch points were used as building blocks of larger systems . The possible makeup of amylopectin unit clusters was investigated via a series of models, including single-single, double-single, and double-double helix systems. The lengths of the single helix section that linked two branch points (internal chains) was systematically varied between values of 0-10 glucose residues. It was found that certain internal chain lengths lead to parallel double helices. It can therefore, be postulated that the length of internal chains may determine the degree of local crystallinity. Furthermore, it was noted that some of the low-energy arrangements of double helices could be superimposed on either the two adjacent and non-adjacent double helices of crystalline A- and B-starch polymorphs
Jump to Vignettes of the representations of the molecular models of amylopectins
In other instances, the distance between the double helices is so large that it may in fact be a model for branching two amylopectin crystals or unit clusters. The results of such a modeling exercise suggest strongly that the branching in amylopectin does not exhibit a random nature . This, in a sense, is not a surprising feature, as the branching process is essentially under enzymatic control. The location and the length of the branching segments, necessarily reflect the three-dimensional structure and the mechanism of the branching enzymes, along with the spatial and temporal availability of the oligosaccharide complexes involved in the synthesis. It might therefore be emphasized that the characteristics of the branching pattern are as equally important as the chain length distribution, playing a determinant role on some physicochemical properties of starch. This work also suggests that chains in the amorphous region may occur in directions different from the main direction of the crystallized chains. Among other, this should explain the flexibility of the amorphous chains during such phenomena as annealing. It may also suggest that the thickness of the amorphous lamellae does not reflect the actual length of the chains in this area, and possibly does not contribute to a certain periodicity in chain length.
The Platelet Nanocrystals�������
The results gathered from the various studies presented above, suggest that amylopectin lamellae are involved as components of complex superhelical architecture. The crystalline lamellae are made of parallel arrays of parallel helices having an average thickness of about 7 nm, with some variations depending on the starch botanical source. The lateral dimensions of these lamellae are less characterized. Starch “micelles” having a diameter of about 15 nm have been reported . The shape and dimensions of the tangential sections of wheat starch granules have been described as hexagonal components with a length of 60 nm and a width of 30 nm . New morphological data on the individual crystalline lamellae of waxy maize starch granules have been reported. By submitting native granules from A-type amylopectin-rich waxy maize to a hydrochloric acid hydrolysis, insoluble residues were obtained . They contained polydispersed platelet nanocrystals that have retained the crystalline A-type of the parent granules. These nanocrystals, that are not totally individualized, have 5 to 7 nm thickness, and characteristic geometrical features such as 60-65° acute angles; they form parallelepipedal blocks with a length of 20-40 nm and a width of 15-30 nm. Analysis of the molecular composition showed a major fraction (73% on a molar basis) of dextrins at DP 6-22 (peak DP at 13) that mostly were linear chains. A minor, branched fraction possessed a peak at DP 26 .
The molecular composition and the thickness of the lamellae is consistent with the presence of double-helical structures made up of single strands having DP between 15 to 18; that would be connected to another chain via α-(1→6)-linkages, in a region that would span about 2.5 nm. The crystalline organization of the double helical components would follow the arrangement found in the A-type structure . From the size of the lateral dimensions of the platelets, one can assess that there is a distinct difference between the morphology of the in vitro grown crystals and these platelet nanocrystals resulting from concomitant biosynthesis and crystallization.
The overall shape of the starch nano-crystals seems to be a homothetic view of the monoclinic unit cell of the A-type allomorph (a = 2.12 nm; b = 1.17 nm). The dimensions of the platelets indicate that between 150 to 300 double-helical components are making up the crystalline domains.. The complementary acute angle of g angle between the a and b axes is 56.5°, which is close to the acute angle of around 65° measured for the platelet crystals. Geometrical considerations taking into account the results of the molecular modelling investigations of duplex formation arising from double helical structures can explain the deviation away from the expected morphology of the platelets. They indicate that within the platelets, morphology is driven by the pairing of parallel double-helices having their axes separated by 1.1 nm, and a relative translation of 0.52 nm. Such an arrangement allows a very strong nesting of the crests and troughs of the double helices, as it expresses complementary shapes complemented by several inter double-helices hydrogen bonds . Based on such considerations, the theoretical construction of the nanocrystals 3D-structures, which is in agreement with the crystal structure of A-type starch, explains the observed morphology characterized by the acute angle around 65°.
a. 
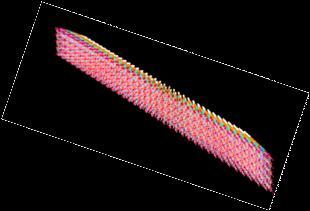
b. 

Fig. 10 Schematic representation of the nanocrystals having (a.) right and (b.) left orientation
These new findings are therefore very relevant to the nascent morphology of starch assembly, indicating that the double helices are not perpendicular to the observation plane, and that the platelet mean plane is inclined with respect to the axis of the double helices. Therefore, these nanocrystals display two main characteristics, i.e. polarity (one crystalline face is different from the other one) and chirality (resulting from the tilt angle between the double-helical axis and the crystalline axis normal to the base plane). The consequence of these features, which can be also extended to B-type starches, in terms of generating super-helical arrangements should be investigated in the context of the results derived from the analysis of the structure of starch granules with small angle X-ray microfocus scattering .

Fig. 11 Starch: The Power of Tenth
Taking all the studies together, we
can be fairly sure about nanoscale
structure (double helices forming
lamellae) and the growth rings
(alternating amorphous and semicrystal
line shells); however, the evidence
for the intermediate structures
(the superhelices and the blocklets) is
less solid. Furthermore, it is still
unclear how the superhelices,
blocklets and growth rings relate to
each other.
Fig. 11 summarises the different structural
levels (glucose units, helices,
lamellae, superhelices, blocklets and
growth rings), from the molecular
(10-9 m) to the microscopic (10-5 m)
level.
